Nach einer Woche Arbeit habe ich meine Zusammenfassung der Zoologie für meine Vordiplomsprüfung abgeschlossen. Ich denke, dass der Inhalt auch anderen Leuten bei ihrer Vordiplomsprüfung weiterhelfen könnte.
Enthalten sind folgende Themen:
Ökologie
Besiedlung eines Lebensraums
Unterschiede zwischen Prädatoren und Parasiten
Logistische Wachstumsgleichung
Ökologische Nische
Autökologie, Synökologie
Symbiosen
Verteilung von Organismen im Lebensraum
Globale Erderwärmung: Anzeichen, Entstehung, Ursachen, wie verhindern, welche Gefahr
Ozonloch: Wirkung, Ursachen, wie verhindern, welche Gefahr
Evolutionsbiologie
Welche Rolle spielen Mutationen bei der Evolution
Mechanismen der Artbildung
Artenvielfalt Tangajikasee, Galapagosinseln, Korallenriff, Antarktis
Prinzipien der phylogenetischen Systematik (Apomorphie, Plesiomorphie)
Entstehung von mittelozeanischen Rücken
Evolutionsraten von Makromolekülen
Hardy-Weinberg-Gesetz
Zellbiologie
Endosymbiontentheorie
Unterschiede zwischen Prokaryonten und Eukaryonten
Querschnitt eines Flagellums
Unterschiede tierische und pflanzliche Zelle
Zoologische Systematik
Insekten – Unterteilung mit Beispielen
Crustaceen
Daphnien, Querschnitt
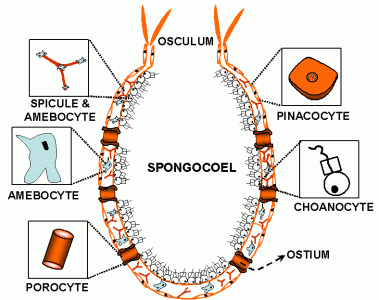
Schwämme – Zelltypen
Coelenterata
Mollusca
Regenwurm – Querschnitt
Stoff- und Energiewechsel
Aerobe Energiegewinnung
Anaerobe Energiegewinnung
Respriratorischer Quotient
Relation Körpergewicht/Energieumsatz
Grundumsatzrate, Aktivitätsumsatz
Ernährung
Herbivore, Omnivore, Carnivore, Saprophage
essentielle Nahrungsstoffe
intra- und extrazelluläre Verdauung
Verdauungstrakte
Verdauungsenzyme der Wirbeltiere
Saures Medium im Magen
Wiederkäuermagen
Atmung/Blut
Kiemen, Schwimmblase
Lungen, Vogellunge
Tracheen
Blutfarbstoffe
Hautatmung
Blutbestandteile
Blutgefäßsysteme
Blutgerinnung
Immunreaktion
Ionen-/Osmoregulation
Ionenpumpen, Ionenkanäle
Osmoregulierer, Osmokonformer
Exkretionsmechanismen, Wirbeltierniere
Thermoregulation
poikilotherme Tiere, homoiotherme Tiere
Thermokonformer, Thermoregulierer
Hormonale Kontrolle
Was sind Hormone
Wichtige Hormone
Steuerungd er Hormonaktivität
Menstruationzyklus
Häutung vom holometabolen Insekt
Pheromone
Download “Zusammenfassung Vordiplom Zoologie” Zusammenfassung-Vordiplom-Zoologie.pdf – 10619-mal heruntergeladen – 704,50 kB